Навигация
Вход в систему

Nothobranchius janpapi Wildekamp, 1977

janpapi: название в честь Jan Pap {Нидерланды }, который открыл этот вид в 1975 и отправил несколько особей любителям в Голландию.
Первое описание
R. H. Wildekamp, 1977 - "Nothobranchius lourensi spec. nov. und Nothobranchius janpapi spec. nov., zwei neue Rivulinen aus Ostafrika" - Das Aquarium, Heft 98, August 1977:326-331; 7 tabs. ;6 figs.
Типовое место обитания
Восточная Танзания, около 80-и км западнее Dar Es Salaam. Впервые пойман Jan Pap в 1975, а затем Jan Lourens в начале июля 1976, в пруду около скоростной дороги из Morogoro в Dar Es Salaam, между мостом через реку Ruvu и деревней Kwaraza.
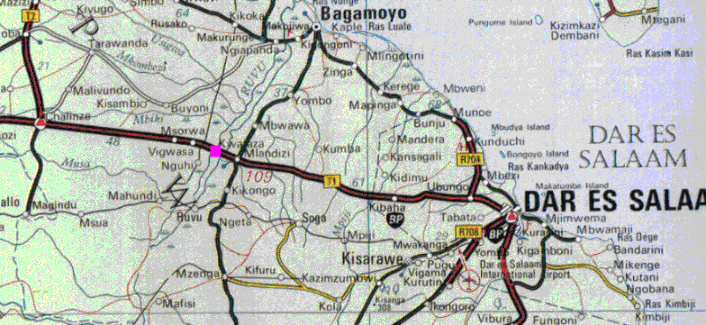
Рис 1: Типовое место обитания N.janpapi около Kwaraza, 80 км западнее Dar-Es-Salaam, на дороге из Dar-Es-Salaam в Morogoro
Wildekamp, (1977) сообщает, что типовое место обитания N.lourensi и N.janpapi являлось прудом около 10 - 15 метров в ширину. Ещё один пруд, расположенный немного дальше, на другой стороне дороги, был около 5 - 7 м в диаметре. Оба пруда сообщались друг с другом под небольшим мостом. 12-ого июля 1976г. глубина обоих прудов была всего около 70 см. Берега были густо заросшими травой и колючим кустарником. В самой воде растительности не было. Температура воды 11-ого июля в 18:15 была 27 С, воздуха - 25 С. рН от 6,5 до 7,0, жёсткость воды не измеряли.
В этой части Танзании сезон дождей продолжается с ноября по май, в январе и феврале количество осадков уменьшается. В 1975 Jan Pap, и в 1976 Jan Lourens наблюдали, что эти пруды полностью пересыхали. N.lourensi и N.janpapi можно найти только в близких водоёмах. Остальные пруды населены N.guentheri, N.melanospilus и неоднолетние рыбы. (Wildekamp, 1977).
Рис 2: Метеорологический профиль Танзании {по данным FAOCLIM}
Таким образом можно предположить что эти пруды питают не только дождевые воды, но так же они должны получать воду и из самой Ruvu River, когда во время длительного дождливого периода река затопляет эту обширную болотистую местность. По данным Wildekamp (1977), присутствие не облигатных однолетних видов рыб в этих водоёмах (таких как Clarias mossambicus Peters, 1852; Alestes affinis Guenther, 1894 and Barbus usambarae (?) Loennberg, 1907) подтверждают это предположение.
Меристические и морфологические данные
Кариотип: n=.
Из первичного описания этого вида, выполненного Wildekamp в 1977, высота тела в 4,1 - 4,9 раза полную длину и в 3,3 - 3,9 раз длину тела. Длина тела составляет 3,7 - 3,9 длинн головы, длина головы равна 1,1 - 1,5 межглазничного гасстояния. Хвостовой стебель в длину в 1,7 - 1,9 раза больше высоты.
Основная форма тела удлинённая, линия спины лишь слегка искривлена. Наибольшая высота тела на уровне брюшных плавников. Спинной плавник сдвинут назад, начинается на уровне 5 или 6-ого луча анального плавника. У обоих полов плавники округлые. Спинной плавник самца удлиннён и заострён сзади. У самцов развёрнутый спинной плавник доходит далеко за начало хвостового стебля, у самок достигает только начала стебля. Рот маленький и направлен вверх.
Под глазом только один ряд чешуй. Расположение чешуй на голове близко к обычному G-типу. На голове пост- и супра-орбитальные канавки органов боковой линии прерывистные, они происходят не от F-чешуй, как у рыб рода Aphyosemion. На верхней губе имеются многочисленные мелкие папиллы. Мелкие папиллы так же имеются на лучах спинного и анального плавника, на спинном только в дистальной части, на анальном - вдоль всей дистальной части.
| Таб 1: Меристические характеристики N.janpapi. (по Wildekamp, 1977 - все измерения приведены в % от длины тела). |
| Характеристика | Голотип | Аллотип | Паратипы |
| Полная длина | 124,9 | 122,8 | 134,8-124,5 |
| Высота тела | 28,2 | 25,3 | 25,1-30,1 |
| Длина головы | 27,0 | 27,3 | 25,5-28,7 |
| Высота хвостового стебля | 13,9 | 10,7 | 10,6-14,9 |
| Длина хвостового стебля | 23,1 | 20,1 | 20,6-20,9 |
| Диаметр глаза | 9,2 | 9,0 | 9,3-9,6 |
| Межглазничное рассточние | 13,6 | 9,3 | 10,8-2,8 |
| Длина рыла | 9,2 | 7,6 | 7,8-8,7 |
| Расстояние спинной плавник-кончик рыла | 64,7 | 69,2 | 69,1-63,6 |
| Расстояние анальный плавник-кончик рыла | 57,6 | 65,1 | 57,3-65,2 |
| Расстояние брюшные плавники-кончик рыла | 42,7 | 48,8 | 42,1-47,2 |
| Характеристика | Голотип | Аллотип | Паратипы |
| спинных лучей | 13 | 12 | 12-14 |
| анальных лучей | 19 | 17 | 17-18 |
| грудных лучей | 16 | 15 | 14-15 |
| чешуй в продольном рыду | 28 | 27 | 26-28 |
| чешуй в вертикальном ряду над началом анального плавника | 8,5 | 8,5 | 8,5 |
| чешуй поперёк хвостового стебля | 5,5 | 5,5 | 5,5 |
| чешуй перед спинным плавником | 17 | 16 | 16-17 |
Существенные морфологические, так же как меристические различия между N.janpapi и морфо-типом Nothobranchius дали возможность Wildekamp (1977) предложить для этого нового вида отдельный подрод, в котором N.janpapi был первым представителем; у взрослых особей N.janpapi расположение чешуй на голове очень близок к G-типу, наблюдаемому у рода Aphyosemion, но без F- и H- чешуй; преорбитальный участок без чешуи набольшой; канавки супра- и посторбитальных органов боковой линии прерывистые; 4 органа боковой линии на рыле погружены в два V-образных канала; у обоих полов на верхней губе большое количество густо расположенных папилл.
В добавок, в сравнении с морфо-типом Nothobranchius, предложенный подрод Aphyobranchius характеризуется меньшим числом лучей в спинном плавнике и расположением спинного плапвника дальше назад. (Wildekamp, 1977).
По описанию N.willerti в 1992 третьего описанного представителя этого подрода после N.luekei (Seegers, 1984), Wildekamp (1992) можно дать пересмотренные определительные характеристики подрода Aphyobranchius: меньший размер; более вытянутая форма тела; сдвинутый назад спинной плавник, прямоугольный анальный плавник у самцов; ктеноидные шипики на чешуе только на нижней части жаберной крышки и ниней части предкрышки; правильное расположение головных чешуй G-типа; предпочтение к поверхностным слоям воды.
Синонимы
Нет
Aphyobranchius является подродом Nothobranchius представлен пелагическими (=предпочитающими поверхностные слои) формами. Все четыре известные на сегодняшний день представители этого подрода включают редко встречающегося N.luekei Seegers, 1984 (до сих под найденного только однажды, 40-км южнее Dar-Es-Salaam, под мостом через Mbezi River, на дороге в Kibiti, Tanzania), N.willerti Wildekamp, 1992 (найденного в системе Lower Tana River, береговые равнины Kenya), N.janpapi Wildekamp, 1977 (Tanzania) и N.sp. "Ifakara" TAN 95/4 (Tanzania).
По данным Wildekamp (1992), N.willerti не придерживается характеристики о предпочтении пелагического обитания в отличие от трёх других представителей подрода и предпочитает держаться в средних и нижних слоях воды. Так же у этого вида не столь хорошо проявляются такие определительные характеристики как удлинённое тело и сдвинутый назад спинной плавник. Wildekamp (1992)таким образом предположил, что Nothobranchius {Aphyobranchius} willerti является наименее специализированным, чем остальные представители подрода и может таким образом считаться наиболее примитивным представителем подрода Aphyobranchius.
N.janpapi и N.luekei являются очень близкородственными видами. Эксперименты по скрещиванию между двумя этими видами не дали потомков. На основании этих результатов Seegers (1987) решил описать новый вид и назвать его N.(Aphyobranchius) luekei, в честь Leuke, который первый успешно развёл этот сложный вид.
По данным Seegers (1984), N.luekei отличается от N.janpapi более удлинённым и сжатым с боков телом, более мелким, чем у N.janpapi размером, ещё более задним расположением спинного и анального плавников, спинным плавником расположенным заметно за анальным отверстием. У N.luekei большее количество лучей в плавниках, чем у N.janpapi, и большее количество чешуй в продольном боковом ряду и перед спинным плавником. В спинном плвнике лучей больше, а в анальном меньше, чем у N.janpapi. Так же есть различия в окраске этих двух видов Seegers (1984).
По данным Van Der Zee (1985), тип хориона в икре N.luekei, сильно отличается от наблюдаемого у N.janpapi и других представителей подрода. Это единственный вид среди всех видов Nothobranchius, у N.luekei наблюдается гексагональный тип, характерный для рода Aphyosemion. На оболочке икринки N.luekei наблюдаются далеко расположенные друг от друга длинные волосоподобные структуры, в то время как у других видов эти структуры намного короче и густо расположены (Van der Zee, 1985).
Популяции
N. janpapi "Kwaraza " TAN 76/?; Tanzania
N. janpapi "Rufiji River Camp" TAN 95/07; Tanzania
N. janpapi "Ruhoi River"; Tanzania
N. janpapi "Ndundu" TAN 95/10; Tanzania
N. janpapi "Bagamoyo" TAN 95/13; Tanzania
N. janpapi "Kwaraza" TAN 95/14; Tanzania
N. janpapi "Kitonga North" TAN 97/9; Tanzania
N. janpapi "Soga" TAN 97/45; Tanzania
N. janpapi "Bagamoyo" TAN 97/39; Tanzania
N. janpapi "Ndundu Ferry" TZ 97/57; Tanzania
N. janpapi "Soga" TAN 98/14 (same as locality TAN 97/45); Tanzania
N. janpapi "Kikongo" TAN 98/09; Tanzania
Голотип
Взрослый самец (33.7 мм полной длины; 21.1 мм длина тела); пойман Rob van Haarlem, Jan Lourens, Theo Steinfort и R. H. Wildekamp 11 и 12 июня 1976, в том же водоёме, что и N.lourensi, на основной дороге между Morogoro и Dar Es Salaam, между мостом через Ruvu River и деревней Kwaraza.
Аллотип
Взрослая самка (35.5 мм полной длины; 28.9 мм длина тела); данные о вылове те же, что и для голотипа.
Паратипы
Один взрослый самец (40 мм полной длины; 32.1 мм длина тела); две взрослые самки (35.2 мм и 35.6 мм полной длины; 28.2 и 28.6 мм длина тела); данные о месте вылова те же, что и для голотипа.
Все взрослые типовые экземпляры содержались живыми 5 месяцев в аквариуме. Были зафиксированы 13-XI-1976 и отданы в Koninklijk Museum voor Midden Afrika, Tervuren, Belgium.
Размер
Самцы могут достигать 50 мм, самки остаются немного мельче, 45мм.
Код
JAN
Распространение и биотоп
Типовое место обитания находится в западной Танзании, примерно в 80-и км западнее Dar Es Salaam, в пруду около основной скоростной дороги из Dar Es Salaam в Morogoro, между мостом через Ruvu river и деревней Kwaraza. Пруд в ширину около 10 - 15 метров. Ещё один пруд, расположенный чуть дальше, на другой стороне дороги около 5 - 7 м в диаметре. Оба пруда сообщаются друг с другом. 12-ого июля 1976г. глубина обоих прудов была около 70-и см. Берега густо покрыты травой и колючими кустарниками. В воде растительности нет. Температура воды 11-ого июня в 18:15 была 27 С, температура воздуха - 25 С, рН между 6,5 и 7,0. (Wildekamp, 1977).
Seegers выловил 3 разных вида Nothobranchius, обитающих симпатрично в одном пруду вдоль Ruhoi River. Это были N.janpapi, N.melanospilus и N.eggersi "Ruhoi" (Seegers, 1985).
В 1995 Wildekamp, Sainthouse и Watters нашли N.janpapi поблизости от Bagamoyo, около переправы через эстуарий Ruvu River, вдоль земляной насыпи. Местность по обеим сторонам насыпи предсталяла собой плоскую травянистую равнину. Густая растительность затрудняла работу сетью. Хотя и не всегда, они находили вместе N.melanospilus и N.janpapi и N.foerschi (Sainthouse, 1996).
В мае 1997 Wildekamp, Watters и Cooper нашли N.janpapi в точке Kitonga North TAN 97/9. В этом месте они обнаружили N.janpapi обитающего симпатрично с N.lourensi и N.fuscotaeniatus.
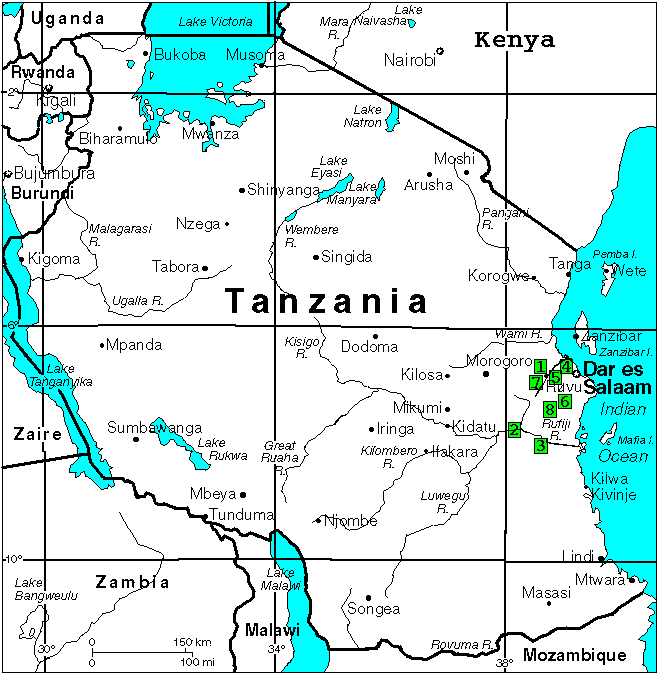
Рис 3: Места вылова N.janpapi в Танзании. 1: "Kwaraza" TAN 76/? и TAN 95/14; 2: "Rufiji River Camp" TAN 95/07; 3: "Ndundu" TAN 95/10 и TZ 97/57; 4: "Bagamoyo" TAN 95/13 и TAN 97/39; 5: "Soga" TAN 97/45 и TAN 98/14; 6: "Kitonga North" TAN 97/9; 7: "Kikongo" TAN 98/09; 8: "Ruhoi River" TZ?/?.
Так же в мае 1997 Wildekamp, Watters и Cooper нашли три разных вида Nothobranchius в месте, обозначенном Soga TAN 97/45. Один из них был Paranothobranchius ocellatus, второй N.janpapi и третий N.foerschi (Watters, June 1998, Killietalk).
В 1997, но в июле, N.janpapi был найден в том же водоёме, где Seegers (1998) впервые обнаружил N.fuscotaeniatus. Seegers (1998) сообщает, что приехав из удачной поездки по Танзании, целью которой был отлов рыб, он решил провести некоторое время в Selous Game Reserve. Так как небольшая переправа в Utete была не в порядке, ему пришлось проехать обратно лишние 60 км до Ndundu, там пересечь Rufiji River, и по более длинной дороге через Kibiti доехать до противоположной стороны реки в Utete, откуда он смог проехать по северному берегу вверх по течению до Selous Game Reserve.
В Nyamwage, дорога на Ndundu резко поворачивает на север. Временами дорога была в очень плохом состоянии, слева и справа располагались рисовые поля и другие залитые водой местности. Вдоль дороги тянулись большие придорожные канавы,вырытые при строительстве дороги. Seegers-у стало ясно, что он приближается к реке и к началу дельты Rufiji River. Так как не представлялось возможным обловить все водоёмы, Seegers решил проверять только некоторые из них. Это дало несколько интересных экземпляров, среди которых были и некоторые Nothobranchius (Seegers, 1998).
Последняя попытка была сделана в остаточном водоёме 2х10 м около дороги (точка TZ 97/57, 2-км южнее переправы в Ndundu через реку Rufiji River). Берега были скрыты высокой травой. В середине былы участки открытой воды. В самой глубокой части уровень воды был 60 см. Под полуденным солнцем Seegers бесценльно закинул сеть и вытащил бедно окрашенную самку Nothobranchius. Так как она не могла относиться к широко распространённому N.melanospilus, он продолжил ловлю. В этот раз он выловил N.janpapi, который его так же мало интересовал, так как тоже широко распространён в этой местности, как и N.melanospilus. В третий раз он выловил самца N.fuscotaeniatus. При полуденном солнце рыба была окрашена в зелёно-голубой цвет с несколькими красно-коричневыми поперечными полосами на теле. (Seegers, 1998).
Нужно было выловлить ещё несколько рыб для того, чтобы найти подходящую самку. Теперь был выловлен самец N.lourensi. Самки этого вида так же были бесцветными, от чего возник вопрос - к какому виду относится первая пойманная самка. Дальше было поймано ещё несколько бесцветных самок, но различий между ними поначалу не было замечено. Наконец Seeger выловил самку, которую никогда раньше не видел, что решило проблему идентификации. Эта самка N.fuscotaeniatus была так же зеленовато-голубая, но не такая яркая как самец. На теле так же имелись различимые красно-коричневые кольцевые линии. Все неокрашенные самки относились к N.lourensi. Заслуживает внимание то обстоятельство, что в этом отрезанном участке придорожной канавы, Seegers не обнаружил никаких других видов рыб, даже широко распротранённого N.melanospilus (Seegers, 1998).
Основываясь на вышеперечисленных метах вылова, территория распространения N.janpapi включает средние и прибрежные части систем рек Ruvu, Ruhoi, Mbezi и Rufiji River. На самом деле эта территория является огромной заболоченной местностью, где рыбы могут свободно распространяться от одного края низменности до другого.
Текущая на севере река Ruvu River имеет свои истоки в горах Uluguru Mountains, около Morogoro. Вначале, она течёт на восток до огромной болотистой низменности, расположенной на востоке Uluguru Mountains. До того как достигнуть этого болота, в неё впадают Rudete и Mgeta Rivers, обе начинающиеся так же в горах Uluguru Mountains. Затем она поворачивает на север, где достигает Индийского океана Indian Ocean, севернее Dar-Es-Salaam, в Bagamoyo.
Рис 4: Основные бассейны рек Танзании (по данным FAO-ALCOM)
Эта болотистая низменность так же связана с большим количеством мелких рек намного большей системы реки Rufiji/Ruaha River, что даёт хорошую возможность для обмена ихтиофауной между системами этих двух рек.
Сходная древняя связь между системами рек Rufiji River и соседними Ruhoi and Mbezi Rivers так же сыграла свою роль в распространении N.janpapi в системах этих рек. Основная территория распространения N.janpapi в восточной Танзании предварительно представлена на рисунке.
N.leukei который близкородственен N.janpapi (как показано выше) до сих пор был выловлен только однажды под мостом через Mbezi River (40 км южнее Dar-Es-Salaam на дороге в Kibiti) Seegers и Wischmann в ходе годовой поездки с 1982 до 1983, хотя три предыдущих раза Seegers ловил рыб в этом водоёме, но безуспешно (Van der Zee, 1985). После поимки N.janpapi около Kitonga (точка: "Kitonga North" TAN 97/9, так же под влиянием Mbezi River) Watters, Wildekamp & Cooper в 1997 стало теоретически вероятным что N.janpapi и N.leukei могут обитать вблизи друг от друга в и вокруг этой речной системы. Так как определния этих двух рыб сходны, они должны быть в состоянии жестокой конкуренции между двумя близкородственными видами так как они никогда не встречаются вместе или должны избегать конкуренции предпочитая разные экологические ниши. Возможное симпатричное обнаружение этих видов в будущем в Mbezi River таким образом будет носить научный характер.
Рис 5: Места вылова и предполагаемая территория распространения N.janpapi в Танзании. 1: "Kwaraza" TAN 76/? и TAN 95/14; 2: "Rufiji River Camp" TAN 95/07; 3: "Ndundu" TAN 95/10 и TZ 97/57; 4: "Bagamoyo" TAN 95/13 и TAN 97/39; 5: "Soga" TAN 97/45 и TAN 98/14; 6: "Kitonga North" TAN 97/9; 7: "Kikongo" TAN 98/09; 8: "Ruhoi River" TZ?/?.
Окраска
Самец: основной цвет тела фиолетово-голубой, коричневатый на спине, на брюхе беловатый, горло жёлтое. На чешуйках имеется тонкая красновато-коричневая кайма. Спинной плавник жёлто-зелёный с несколькими коричневыми крапинками, у основания переходит в серо-голубой. В дистальной части имеется тонкая беловато-голубая кайма. Хвостовой плавник - желтовато-зелёный, в проксимальной части сверху и снизу красные штрихи. В основании анальный плавник жёлтый, переходящий в ярко-оранжевый. Грудные планики бесцветные с небольшой светло-голубой каймой. Ирис глаза голубой.
Самки: Серо-коричневые, брюшко и ирис глаза серебристые; все плавники бесцветные.
Содержание и разведение
N.{Aphyobranchius} janpapi являются атипичным для Nothobranchius, так как населяют верхние слои воды предпочтительнее, чем нижние. Мальков, так же как (суб)взрослых особей можно часто наблюдать за поиском корма около поверхности. В естественных водоёмах взрослые особи предпостительно держатся открытых пелагических зон между затопленными кутами и ветками.
Этот вид можно разводить в овде со следующими характеристиками: рН 6,8 - 7,0, жёткость воды между 9 и 16 и температуре 23 - 25°C (73 and 75°F). Такую воду легко получить добавив дождевую к обычной водопроводной воде. При слишком высокой температуре водержания существенно уменьшает продолжительность жизни. Добавление соли (1 чайная ложка на 10 литров воды) как превинтивная мера против оодиниоза.
Аквариум для содержания лучше густо засадить водным папоротником или подобными растениямидля обеспечения самкам и слабым самцам места для укрытия. Частая подмена воды так же необходима для хорошего самочувствия этих рыб.
Самцы не агрессивны по отношению друг к другу. Более сильные самцы всё же вытесняют более слабых на более освещённые участки аквариума. Если аквариум достаточно просторный, порванные плавники и побитые рыбы наблюдаются редко. В просторном аквариуме можно содержать одновременно несколько самцов одновременно, если на каждого приходится 10 - 15 литров воды. Нужно избегать посадки 2-х самцов в один маленький аквариум; в этом случае более сильный самец буде почти постоянно преследовать более слабого или даже убьёт его.
Для разведения можно использовать тройку рыб, но рекомендуется сажать несколько самок на одного самца (3 - 4 самки) и сажать на нерест сразу группу особей.
Особенным для этого вида является тот факт, что производители не опускаются ко дну для того, чтобы отложить икру. Нерест может происходить в любом месте аквариума, предпочтительнее на открытых участках в средних слоях воды. Таким образом этот вид не нуждается слое торфа. Зато можно исползовать любой субстрат, покрывающий как можно большую поверхность дна нерестовика. Это нужно не для концентрации икры, а для того, чтобы сразу "поймать" её. Моно экспериментировать с торфом. Часто производители откладывают икру в и на торфяной субстрат охотнее, чем на голое дно аквариума.
2 - 5-и мм слой торфа в открытом пластиковом "корыте"можно использовать как место для нереста. В таком нерестовике рыб оставляют на нересте 5 - 7 дней, после чего торф удаляют, аккуратно отжимают и отавляют подсыхать до хорошо известного состояния табака (верхний слой становится светло-коричневым). Затем нерестовый субсбтрат с икрой помещают в закрытый пластиковый пакет с достатоным количеством воздуха. Икра N.janpapi хранится 2,5 месяца при постоянной температуре около 24 °C (74°F). Я получал успешный выклев даже после 7-и месячного сухого периода у популяции TAN 97/9 "Kitonga North".
Выклев развившихся мальков происходит в обычной отстоянной (1 неделю) прохладной водопроводной воде (18°C or 64°F). Водой заливают торф и встряхивают, чтобы икра смочилась и опутилась на дно. Затем торф всё ещё плавающиё на поверхности удаляют, чтобы только что выклюнувшиеся слабые мальки имели более свободный доступ к поверхности (для заполнения плавательного пузыря).
В аквариуме, где происходит выклев, объём которого должен быть не менее 5-и л, икра должна быть закрыта слое воды 20 - 50 мм. Иногда для выклева первого малька требуется 24 часа, последние выклёвываются в течение нескольких дней.
Так как только что выклюнувшийся малёк очень мелкий (1,5 мм длиной, мельче чем малёк N.patrizii или N.rachovii) им нужно давать "зелёную воду" с большим количеством инфузории и микроорганизмов. Даже самый крупный малёк не в состоянии сразу поедать даже самых мелких науплиусов Артемии. До того, как малёк N.janpapi сможет брать только что выклюнувшихся науплиусов Артемии он должен оставаться 2 недели в "зелёной воде". Поэтому, перед тем как заливать икру нужно убедиться в наличии богатой культуры инфузории или "лелёной воды" во время выклева. Если богатая микроорганизмами вода во время выклева не готова - её, как правило, невозможно быстро приготовить и разведение этого вида ставиться под угрозу. Поэтому полезно постоянно держать в саду или на балконе ёмкость со старой водой и опавшими листьями. Через кокое-то время эта среда будет богата необходимыми микроорганизмами для любого мелкого малька.
Всё же лучше использовать культуру крупной инфузории Paramecium, которую немецкие любители нашли в Камеруне в начале 90-х годов. Эта чистая культура Paramecium, лишённая запаха, легко поддерживается круглый год в обычной отстоянной водопроводной воде при добавлении 1 - 2 капель молока раз в неделю на 5-и литровую банку культуры (не перекармливайте культуру, иначе она будет плохо пахнуть). В этом случае под рукой всегда будет культура инфузории, пригодная для кормления самого мелкого малька. Когда количество корма в аквариуме с мальками уменьшается, 10-20 мл культуры инфузории добавляют прямо в аквариум с мальком. Используя это корм ускорит рост мальков до недели.
Для выклева я использую аквариум на 2 литра. Как только малёк начинает свободно плавать, я переношу его в болший аквариум, содержащий "зелёную воду" из черепашьего пруда моей дочери, доведённую нужной температуры или выше упомянутую культуру инфузории. Больший размер аквариума кроме того, что обеспечивает более стабильную среду для роста малька, там меньше откладывается продуктов жизнедеятельности и обеспечивает бюольшее количество микроорганизмов. Я так же обычно добавляю один или два размельчённых сухих листа. К тому же, большой аквариум делает не столь необходимой частую подмену воды и позволяет регулярно повышать уровень воды не беспокоя при этом малька.
Использование большого стабилизированного аквариуме обеспечивает защитную среду для слабого малька. Присутствие несколько часов в день яркого верхнего света нужно для того, чтобы мальки могли найти движущийся корм. При таком оборудовании аквариума нет никакой необходимости в фильтрации или даже небольшом движении поверхностного слоя воды. Большинство микроорганизмов собираются непосредственно под поверхностью воды, образуя тёмный плавающий слой, который не следует удалять. В этом слое мальки ищут корм.
Примерно после 1 - 2 недель, мальки начинают поедать самых мелких только что выклюнувшихся науплиусов Артемии, теперь большинство проблемм позади. Всё же это довольно медленно растущий вид и скорость роста, наблюдаемая у других Nothobranchius, никогда не достигается у рыб подрода Aphyobranchius. Примерно через 3 - 4 месяца тервые самцы начинают окрашиваться, а активными становяться к 5 - 6 месяцам.
Они являются значительным добавлением для аквариума, населённого только Nothobranchius, так как они занимают верхний и средний слои воды, подобно рыбам рода Aphyosemion.
Литература
© Авторское право 2000, Marc Bellemans. Все права зарезервированы. Оригинал статьи
- Войдите на сайт для отправки комментариев
Новые обсуждения форума
- Группы в месенджерах
- Выставки Killi в 2025 году (РЕЗУЛЬТАТЫ)
- Предновогодняя встреча 2024.
- Выставки Killi в 2024 году (РЕЗУЛЬТАТЫ)
- Февральская встреча 9 числа
- Поздравляем!
- Астролебиасы (Austrolebias)
- Лягушатники - тоже люди?
- Выставки Killi в 2023 году (РЕЗУЛЬТАТЫ)
- Sergejam Dzim?anas Diena!
- "Моно" аквариум самосевом.
Активные обсуждения форума
- Продам эпиплатис Дагета .
- Группы в месенджерах
- Выставки Killi в 2025 году (РЕЗУЛЬТАТЫ)
- Предновогодняя встреча 2024.
- "Моно" аквариум самосевом.
- Выставки Killi в 2024 году (РЕЗУЛЬТАТЫ)
- Общая выставка с нашим участием.
- Февральская встреча 9 числа
- Сведения о биотопах
- Александр Сухов с днём рождения!
- Поздравляем!
Новые записи в блогах
- Всем привет. Немного психотерапии для нашего сообщества
- Epiplatys dageti
- Один из способов фотографирования килли
- Fundulopanchax walkeri spurelli
- N. Rachovii
- Aphyosemion australe фото дня
- Fundulopanchax gardneri "Gold" фото дня
- Аквариумные Аплохейлусы
- Разведение Fundulopanchax gardneri nigerianus
- Про выставку аквариумных рыб